Эта записка является текстом доклада, подготовленного для семинара в Московском институте информационных технологий. Слайды можно посмотреть здесь.
 |
| Рис.1. Жизнь = Химия + Кибернетика. |
Определения
Перед тем, как начать наш разговор, думаю, стоит сказать несколько слов о предмете предстоящего обсуждения и привести некоторые полезные определения. Вопросы возникновения и поддержания жизни на Земле, которых мы коснемся с научной стороны, по самой природе своей выходят на вопросы философско-религиозные (эти последние мы только упомянем и в дальнейшем касаться не будем). Именно поэтому осмысление начал биологической науки — в том числе, с точки зрения информации — приводит к довольно накаленным дискуссиям между подчас непримиримыми оппонентами. Мы, надеюсь, постараемся удержаться в рамках научной полемики. Я ни в коем случае не принуждаю никого разделять мою точку зрения на то, что жизнь представляет собой артефакт и теперь поддерживается в биосистемах благодаря встроенным в нее механизмам репликации и развития дочернего организма или, говоря языком религии, что жизнь была однажды сотворена. Я просто буду приводить аргументы в пользу моей позиции, по возможности не пересекая демаркационную линию и оставаясь в рамках науки.
По моему глубокому убеждению, в основе научной деятельности в первую очередь лежат особое чутье и умение задавать нужные вопросы, и только во вторую находить способы их решения. Мы с вами, дорогие слушатели, являемся счастливыми свидетелями того, как наука о жизни переживает один из интереснейших периодов своего развития, когда по законам диалектики накопившиеся на сегодняшний день научные данные уже не укладываются в рамки представлений, устоявшихся за прошедшие с момента появления классической теории эволюции полтора столетия. Следить за выработкой новых идей и теорий – процесс поистине увлекательный. И я считал бы свою задачу выполненной, если бы хотя бы отчасти мне удалось зажечь уважаемую аудиторию интересом к обсуждаемым вопросам.
Итак, определения.
Управлением мы будем называть совокупность действий лиц, принимающих решения (ЛПР) по переводу произвольной системы в целевое состояние, характеризующееся оптимумом некоторой функции цели. Результат этих действий может быть представлен системой управления.
Сложностью по Колмогорову некоторого объекта будем называть наименьшую возможную длину программы его описания без потери информации на некотором универсальном языке.
Системы в хаотическом состоянии характеризуются высокой степенью энтропии и неопределенности и, следовательно, высокой сложностью, что видно по знаменитой формуле Шеннона информационной двоичной энтропии для независимых случайных событий x с n возможными состояниями (p — функция вероятности):

- Действительно, чем меньше вероятность реализации определенного исхода, при котором случайная переменная принимает определенное значение из n возможных, тем больше информации получает наблюдатель в случае, когда этот исход реализуется. С другой стороны, максимального значения энтропия достигает при условии, что каждое из n состояний равновероятно (хаос). Длина представления значения некоторой переменной y при использовании символов булева алфавита {0,1} равна округленному до большего целого log2y. Поэтому для передачи информации о состоянии одной случайной величины, допускающей два значения, требуется максимум 1 бит информации (рис.2).
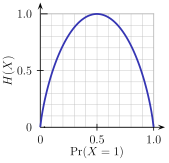 |
| Рис.2. Информационная энтропия (в битах) случайной величины Х, принимающей два значения (например, результат бросания монеты: решка Х=0 и орел Х=1). По горизонтали отложена вероятность того, что X принимает значение 1 (выпадает орел). Энтропия принимает максимальное значение 1 бит в случае равной вероятности результата ("честный" бросок монеты). Энтропия равна 0 в случае, если тот или другой результат имеют максимальную вероятность 1.0 (если к противоположной стороне монеты прилеплен грузик). Источник: Википедия, статья Entropy (Information Theory). |
Напротив, системы в упорядоченном состоянии, например, кристаллы, характеризуются меньшей неопределенностью, что предполагает существование более коротких программ описания вследствие возможности информационного сжатия. Интересно заметить, что живые системы отстроены по сложности всего на несколько процентов от хаотического состояния. Это, по всей видимости, вдохновляет многих сторонников теорий самоорганизации материи. Но дело в том, что функциональность не зависит напрямую от степени сложности системы. Да, действительно, биосистемы находятся ближе к хаосу с точки зрения сложности, чем к упорядоченному состоянию, но хаос/порядок и функция — совершенно разные вещи. В связи с этим следует с сожалением отметить некоторую путаницу в устоявшихся определениях. То, что обычно в литературе именуется самоорганизацией, по сути является лишь самоупорядочением. Давайте коснемся этого вопроса подробнее.
Организация предполагает существование иерархии и формального управления в сложных системах, тогда как упорядочение подразумевает генерацию некоторых относительно простых конфигураций. Современная наука не располагает никакими данными, которые бы подтверждали возможность самопроизвольного возникновения управления и организации как таковой. Наблюдается лишь спонтанное упорядочение.
Иными словами, экспериментальное подтверждение находит лишь свойство материи спонтанно порождать низкоинформационные структуры фрактального типа. Эти конфигурации регулярны, монотонны, избыточны, просты по Колмогорову и, как мы видели выше, поскольку несут в себе малое количество информации, достаточно плотно сжимаемы.
Примерами таких структур являются кристаллические решетки, конвекционные паттерны (так называемые ячейки Бенара), интерференционные картины, рисунки песчаных дюн. К процессам, приводящим к подобным структурам, следует также отнести автокаталитические циклы, химические часы и пр. Но как только мы имеем дело с формальным управлением в системах, по имеющимся в большом количестве научным данным мы необходимо приходим к выводу об искусственном происхождении таких систем. Экспериментальная наука говорит нам о том, что за формализмом всегда стоит интеллект. Это не учитывают имеющиеся теории самоорганизации типа кауффмановского антихаоса, айгеновых гиперциклов или пригожинских диссипативных структур. Знаменитая S-теорема Климонтовича [Климонтович 1996], на которую обычно ссылаются сторонники абиогенеза и тому подобных гипотез, действительно устанавливает возможность спонтанного понижения энтропии в открытых системах, находящихся достаточно далеко от динамического равновесия. Но это еще не означает возможности спонтанного появления иерархических связей между компонентами системы и «кристаллизации» формального управления.
Под функциональностью мы будем понимать возможность произвольной системы достичь достаточно близких к оптимуму значений функции цели, или обобщенной полезности совместного действия множества компонентов как единого целого. В применении к живым организмам мы говорим о биофункциональности, включающей в себя поддержание гомеостаза (в чем исключительно важную роль играет обмен веществ), а также размножение и реагирование на стимулы.
Примерами таких структур являются кристаллические решетки, конвекционные паттерны (так называемые ячейки Бенара), интерференционные картины, рисунки песчаных дюн. К процессам, приводящим к подобным структурам, следует также отнести автокаталитические циклы, химические часы и пр. Но как только мы имеем дело с формальным управлением в системах, по имеющимся в большом количестве научным данным мы необходимо приходим к выводу об искусственном происхождении таких систем. Экспериментальная наука говорит нам о том, что за формализмом всегда стоит интеллект. Это не учитывают имеющиеся теории самоорганизации типа кауффмановского антихаоса, айгеновых гиперциклов или пригожинских диссипативных структур. Знаменитая S-теорема Климонтовича [Климонтович 1996], на которую обычно ссылаются сторонники абиогенеза и тому подобных гипотез, действительно устанавливает возможность спонтанного понижения энтропии в открытых системах, находящихся достаточно далеко от динамического равновесия. Но это еще не означает возможности спонтанного появления иерархических связей между компонентами системы и «кристаллизации» формального управления.
Под функциональностью мы будем понимать возможность произвольной системы достичь достаточно близких к оптимуму значений функции цели, или обобщенной полезности совместного действия множества компонентов как единого целого. В применении к живым организмам мы говорим о биофункциональности, включающей в себя поддержание гомеостаза (в чем исключительно важную роль играет обмен веществ), а также размножение и реагирование на стимулы.
Кибернетическое сечение: ограничения и управление
Представим себе некоторое пространство S, состоящее из двух подпространств Sогр и Sупр, так что S = Sогр + Sупр. Sогр содержит системы, в которых реализуются физико-химические ограничения. Подпространство Sупр содержит множество управляющих функций, то есть концептуальные схемы формального управления кибернетическими системами, функционирующими по произвольно задаваемым правилам (рис.3) для достижения определенной цели. В то время как ограничения, присутствующие в любой реальной физической системе, определяются теми или иными физико-химическими взаимодействиями в каждом конкретном случае, добавление в систему управления соответствует осуществлению осознанного выбора в рамках определенного формализма и на практике всегда производится целенаправленно интеллектуальным агентом (ЛПР). Здесь под осознанным выбором следует понимать особую, третью Аристотелеву категорию причинности, отличную от случайности и закономерности. Поясню, что я имею в виду.
 |
| Рис.3. Некоторые примеры систем, функционирование которых определяется формальными правилами. В таких системах возможна реализация творческого замысла. |
Понятно, что управление, также как и ограничения, на практике неизбежно представлено материально, или "загружено в физическую реальность", по выражению Дэвида Абеля. Однако физическая реальность сама по себе может предоставить лишь субстрат для реализации управления. Управление же, реализуемое поверх физико-химического уровня, не сводится лишь к физико-химическим взаимодействиям. Именно поэтому мы и говорим об особом подпространстве формального управления Sупр.
Эмпирическая наука говорит нам, что в пространстве S возможен лишь переход из Sупр в Sогр, но не наоборот. Указанный переход реализуется созданием интегральной схемы управления, состоящей из логических элементов — настраиваемых переключателей, несущих по 1 биту информации и актуализирующих целенаправленный выбор со стороны ЛПР по достижению какой-либо цели. Состояние каждого отдельного переключателя не зависит от физико-химических ограничений, но выбирается ЛПР по определенным заданным правилам с целью оптимизации целевой функции. Схема, набранная из переключателей, таким образом, представляет собой своеобразный мост из Sупр в Sогр. Заметим, что ЛПР решает, какие именно значения будут принимать булевы переменные в каждом переключателе для оптимизации заданной функции цели. Это назначение не является результатом действия каких-либо физических закономерностей или случайности, поскольку процесс назначения от них не зависит. Переменные получают именно такие, а не иные значения лишь в результате осознанного выбора конструктора/проектировщика. Таким образом, отсюда видно, что поскольку организация интегральной схемы управления не зависит от ограничений, но диктуется правилами, с формальной точки зрения организация управления соответствует реализации осознанного выбора ЛПР.
Итак, управление не сводится лишь к физическим процессам, реализующим ограничения. Интуитивно это совершенно ясно, поскольку смысл сообщения не
сводится лишь к реализации физического процесса его передачи. Ведь для восприятия информации, передаваемой, скажем, по компьютерной сети, недостаточно рассмотреть лишь скачки напряжения на физическом уровне стэка TCP/IP, но необходимо иметь представление о том, как эту информацию декодировать для получателя. Декодирование же производится по соглашению (протоколу) и следует правилам, не зависящим от процессов физического уровня (подробнее см. сетевую модель OSI, о чем мы скажем ниже). В отличие от ограничений, правила с их семантикой задаются произвольным образом поверх физического уровня. Например, при действиях с булевым алфавитом {0,1} мы вольны поставить в соответствие единице значение НЕТ, а нулю — ДА. Правила легко могут меняться нами в зависимости от контекста.
Повторим, что эмпирическая наука предоставляет нам примеры перехода из Sупр в Sогр, но не наоборот. Интегральные схемы, задающие формальные предписания для реализации развития, развертывания, онтогенеза кибернетических систем, на практике не возникают спонтанно из хаоса. По замечанию Дэвида Абеля, массив данных, подтверждающих исключительно однонаправленный характер данного перехода настолько значителен, что указанной однонаправленности должен быть присвоен статус закона природы. Действительно, неживая природа в принципе инертна по отношению к формальным процессам, в том числе к таким, как оптимизация функции, организация кибернетического управления и др., функционируя в рамках физических ограничений, тогда как формализм оперирует правилами. На базовом уровне физической реальности ни управления, ни целей не существует. Модели, подобные кауфмановской границе хаоса, предполагающие гипотетическую спонтанную огранизацию управления на физическом уровне, не находят подтверждения в реальности. Жизнь не может самоорганизоваться подобно кристаллам, поскольку спонтанная кристаллизация из раствора является не чем иным, как действием лишь физических органичений, а не управления в строгом смысле слова. Доказательством этому служит низкое информационное наполнение регулярных структур и, как следствие, подверженность таких структур высокой степени информационного сжатия, в
то время, как живые системы изначально функционально сложны и характеризуются наличием кибернетического управления и скоординированных формальных процессов.
Итак, сообщение системе функциональности и управления, наделение информационного обмена смысловой нагрузкой, а также интегрирование различных подсистем в единое устойчиво функционирующее целое представляет собой формальный процесс или совокупность процессов. Организация всякого формального процесса требует реализации целенаправленного выбора, во всех без исключения случаях на практике осуществляющегося осознанной деятельностью ЛПР.
Контекст функционирования биосистем
Чем же отличается биохимия от химии? Являются ли ДНК/РНК "просто молекулами" или они несут на себе следы интеллектуального воздействия? Биохимия отличается своим контекстом и семантикой, смысловой нагрузкой генетической информации, представляющей собой не что иное, как шаблон для сложнейших массово-параллельных процессов синтеза биологических структур дочернего организма. В первом приближении развертывание биохимических процессов включает два уровня:
- нижний уровень А непосредственных физико-химических взаимодействий;
- верхний уровень B управления физико-химическими взаимодействиями.
Имеющуюся картину можно сравнить с системой представления информации в сетевой модели OSI, представляющей собой стэк (иерархию) протоколов, использующихся при
организации информационных потоков в компьютерной сети. Уровень А будет соответствовать физическому уровню стэка OSI (рис.4), уровень B – прикладному уровню, то есть самому верхнему уровню, доступному пользователям различных приложений.
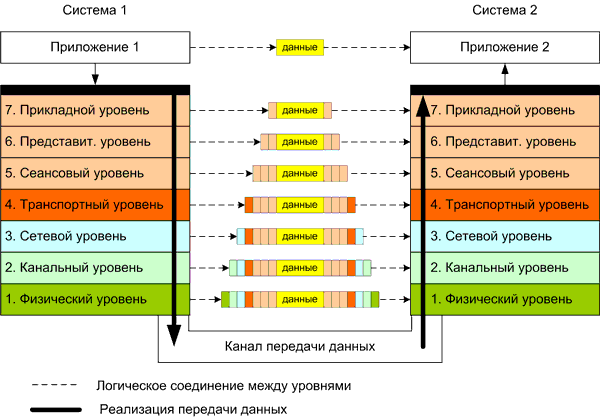 |
| Рис.4. Организация передачи данных согласно модели OSI между двумя приложениями, выполняющимися на подключенных к сети устройствах. Источник: IndustrialNets.ru. |
Процесс обработки информации в персональном компьютере или ином устройстве отправителя идет от прикладного уровня вниз до физического, на котором
посылаемые данные преобразуются в биты и, в конечном счете, в скачки напряжения в сетевом кабеле. Процесс обработки информации в компьютере (устройстве) получателя обратный: скачки напряжения детектируются, преобразуются в поток данных, передаваемых с уровня на уровень вверх до прикладного, на котором они отображаются с помощью конкретного используемого приложения/программы в виде, удобном для чтения человеком.
Например, Вы указываете в браузере веб-адрес (URL) интересующей Вас страницы в интернете. Браузер обрабатывает Ваш запрос, преобразуя URL в сетевой адрес сервера, посылает запрос этому серверу, получает отклик от него в виде пакета запрашиваемых данных, например, в формате HTML (при условии, что адрес правильный и сервер подключен к сети в данный момент) и, наконец, обрабатывает HTML-данные и отображает содержимое страницы в Вашем браузере.
Именно на уровне приложений задается семантика будущего обмена информацией в сети. Скажем, автор статьи набивает текст, пользуясь текстовым редактором, и затем публикует ее на сервере. Для успешного прочтения статьи, она должна находиться по указанному адресу в формате, "известном" Вашему приложению (в данном случае, браузеру).
При этом важно отметить несколько обстоятельств, существенных с точки зрения организации успешного обмена информацией.
- Для корректной интерпретации записанной на носителе информации необходимо предварительно обеспечить совместимость/совпадение протоколов ее представления/считывания, используемых отправителем и получателем (рис.5).
- Представление информации на разных уровнях имеет формальный характер. Протокол есть не что иное, как набор правил, наложенных поверх физико-химических взаимодействий; в свою очередь, физико-химические взаимодействия определяют лишь ограничения, в которых функционирует данная система, но не сами указанные правила.
- Поэтому правила как таковые несводимы к физическим закономерностям, задающим ограничения работы системы (как и правила высших уровней несводимы к правилам низших).
- Наличие правил достоверно указывает на интеллектуальную деятельность проектировщика/разработчика, поскольку сама физическая реальность функционирует лишь в рамках законов, будучи инертной по отношению к правилам.
 |
| Рис.5. Передача информации от источника получателю по физическому каналу с использованием общего протокола представления. |
То же самое происходит в живой клетке, с разницей в конкретных деталях, а именно: в реализации протоколов записи/считывания. Физический уровень (уровень А) представлен биополимерами: молекулами ДНК/РНК, несущими нуклеотиды четырех типов: A, C, T (U в РНК), G.
Триплеты нуклеотидов (кодоны, рис.6) интерпретируются считывающей полимеразой и в случае белок-кодирующей части молекулы соответствуют, как правило, отдельному аминокислотному остатку, который впоследствии будет синтезирован как часть молекулы белка. Запись/cчитывание инициируется и завершается особыми старт-стоп кодонами. Свойства синтезируемых белков определяются последовательностями аминокислотных остатков. Именно этими последовательностями и представлен прикладной уровень (уровень В), то есть не чем иным, как биологическим содержанием генетической информации.
Итак, мы видим, что запись/cчитывание генетической информации есть формальный процесс, алгоритм, или последовательность инструкций, заданных с помощью системы материальных символов и записанных на биополимерном носителе. Современная наука не знает ни одного примера самопроизвольной генерации формальных предписаний, протокола их обработки, а также загрузки этого протокола в систему. Напротив, наличие этих признаков есть свойство лишь артефактных систем обработки информации.
Обстоятельства, которые мы отмечали в связи с обсуждением стэка протоколов сетевой модели OSI, справедливы и в отношении живой клетки. Существование априорно установленного протокола (набора правил) кодирования/интерпретации генетических инструкций необходимо точно так же, как необходимо существование подобных протоколов и в искусственных системах обработки информации. Таким образом, жизнь некорректно сводить лишь к химии, если, конечно, мы хотим получить адекватное представление о том, что такое жизнь с точки зрения современной науки.
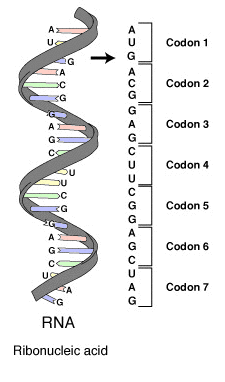 |
| Рис.6. Серия кодонов части молекулы информационной (матричной) РНК (mRNA). Каждый кодон состоит из трех нуклеотидов, обычно соответствующих отдельной аминокислоте. Нуклеотиды именуются первыми буквами их химических соединений: A (adenine), U (uracil), G (guanine) и C (cytosine). В состав данной молекулы mRNA входит U (uracil), вместо которого в ДНК присутствует T (thymine). Молекула mRNA впоследствии инструктирует рибосому синтезировать белковую молекулу согласно последовательности кодонов. Источник: Википедия, статья Genetic code (перевод наш – Е.С). |
Отмечу, что нет нужды представлять то, о чем я говорю, как попытку наделения носителей генетической информации самих по себе мистическими свойствами. Напротив, свойства вполне химические, но только лишь химией не исчерпывающиеся. Мы выплескиваем вместе с водой и ребенка, когда сводим сложнейший семиотический процесс, действительно имеющий химическую составляющую, только лишь к этой составляющей. Можно, конечно, утверждать, что страница газеты содержит всего-навсего причудливо упорядоченные пятна краски. На физическом уровне так оно и есть. Однако этим не исчерпывается содержание газетной публикации. Точно так же смысл человеческой речи нельзя свести исключительно к звуковым колебаниям, а функционирование сложных приборов, таких как, например, телевизор или радиоприемник, не может быть объяснено только лишь существованием электрического тока.
Как я уже говорил, семантическая нагрузка сообщения во всех без исключения примерах функционирования искусственных систем обработки информации есть следствие осмысленных действий лиц, принимающих решения. Таким образом, распознавание семантической нагрузки достоверно указывает на интеллектуальную деятельность проектировщика кибернетической системы. Семантика сообщается той или иной конфигурации материи лишь по выбору ЛПР (волевым актом).
Это особенно видно на примере шифров и условных знаков, так хорошо известных из приключенческих кинофильмов. Скажем, расставленные определенным образом предметы могут служить сигналом к какому-либо действию. Вспомним хотя бы горшок с цветами на окне как сигнал о провале советского разведчика в Берне из киноленты "17 мгновений весны". При этом акторы, не имеющие доступа к ключу шифра, будут интерпретировать данную конфигурацию лишь шумом. Офицеры гестапо, задержав нашего резидента, естественно, ничего не заподозрив, не тронули цветы на подоконнике.
Это особенно видно на примере шифров и условных знаков, так хорошо известных из приключенческих кинофильмов. Скажем, расставленные определенным образом предметы могут служить сигналом к какому-либо действию. Вспомним хотя бы горшок с цветами на окне как сигнал о провале советского разведчика в Берне из киноленты "17 мгновений весны". При этом акторы, не имеющие доступа к ключу шифра, будут интерпретировать данную конфигурацию лишь шумом. Офицеры гестапо, задержав нашего резидента, естественно, ничего не заподозрив, не тронули цветы на подоконнике.
Мне кажется, для прояснения сказанного может быть полезной следующая аналогия. Рассмотрим классы сложности различных задач разрешимости (decision problems).
- На мой взгляд, эта аналогия служит и хорошей иллюстрацией понятия нередуцируемой сложности (irreducible complexity), о котором много спорят биологи. Это понятие было впервые предложено Майклом Бихи (Michael Behe) в книге "Черный ящик Дарвина" в 90-е гг. прошлого века. Впрочем, споры касаются вопросов происхождения неустранимо сложных биосистем, а не самого понятия. Но эту интересную тему я не буду здесь затрагивать.
Под сложностью алгоритма решения задачи будем понимать число шагов его выполнения. Соответственно, сложность задачи есть число шагов наиболее эффективного алгоритма ее решения. Будем называть быстрым алгоритм решения задачи разрешимости, приводящий к ответу "ДА/НЕТ", за число шагов, не большее полинома от размера N исходных данных. На рис.7 изображены классы сложности задач разрешимости. Известно, что каждый нижестоящий дочерний класс сложности вложен в вышестоящий класс, что отражено ребрами графа, представленного на рисунке. Однако вопрос о вложенности вышестоящих классов в нижестоящие до сих пор является открытым. Вполне вероятно - и с этим согласно большинство специалистов по теории сложности, - что вышестоящие классы вообще не сводятся к дочерним классам преобразованиями полиномиальной сложности от N. Еще раз повторим, что взаимная сводимость двух каких-либо классов сложности на рис.7 эквивалентна равенству этих классов и является открытой математической проблемой. Например, проблема доказательства неравенства классов P и NP объявлена институтом Клэя одной из пока не решенных задач тысячелетия.
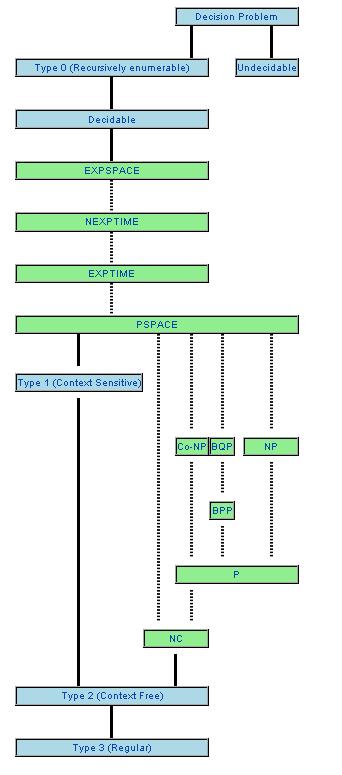 |
| Рис.7. Классы сложности задач разрешимости. Источник: Wikipedia, Complexity Class. |
- NP (nondeterministically polynomially solvable) - класс задач комбинаторной сложности, (то есть задач, для которых не найдено быстрых алгоритмов), но все же допускающих быструю проверку корректности предполагаемых решений (т.наз. сертификатов). Задачи класса NP быстро решаемы на недетерминированной машине Тьюринга, включающей оракул. Пример задачи из класса NP - нахождение кратчайшего маршрута через M точек на плоскости (знаменитая задача о коммивояжере). Вполне очевидно, что проверить имеющийся маршрут, на предмет того, включает ли он все M точек и каждую только один раз, можно за O(M) шагов, то есть достаточно быстро.
- P (polynomially solvable) - класс задач, допускающих быстрые алгоритмы решения. Примерами задач из класса P являются упорядочение массива чисел и нахождение решения системы алгебраических уравнений на множестве действительных чисел. Известно, что решение системы из n уравнений можно отыскать за O(n2) шагов с помощью метода Гаусса исключения переменных.
- Заметим, что требование целочисленности решений переводит задачу в класс NP (см. диофантовы уравнения).
Задачу из класса P всегда можно представить как задачу из класса NP. Действительно, тот или иной известный быстрый алгоритм решения исходной
задачи может выступить в роли оракула в недетерминированной машине. Однако, неизвестно ни одного способа сведения NP-сложной задачи к задаче из класса Р в общем случае.
ДНК как предписание. Фундаментальные проблемы макроэволюции по Дарвину
Известно, что ДНК представляет собой запись онтогенетической информации, или предписание того, как должно протекать развитие биосистемы. Эта информация по природе функциональна, а ее запись и считывание представляют собой формальные процессы (алгоритмы). Иными словами, для записи, считывания и интерпретации информации в генотипе существует определенный язык со своим алфавитом, синтаксисом и семантикой. Поэтому ДНК есть код в строгом смысле слова. Однако, единственным известным эмпирической науке на сегодня источником формализма является интеллект.
Таким образом, предположение о самопроизвольном появлении высших таксонов (классов, типов) дарвиновским градуалистическим способом равносильно предположению о возможности спонтанной генерации принципиально новых формальных инструкций в достаточно больших количествах. Какие-либо экспериментальные данные, подтверждающие эту гипотезу, совершенно отсутствуют.
Несмотря на то, что наука располагает данными, позволяющими предполагать возможность чрезвычайно высокой скорости видообразования, ведущих к формированию низших таксонов (как например, в опытах Г.Х. Шапошникова с тлей), тем не менее, основной массив данных подтверждает лишь случайный характер мутаций уже имеющихся таксономических единиц (см., например, результаты длительных экспериментов с бактериями E-coli). То есть ограничения, накладываемые дарвиновским отбором, вторичны по отношению к существующему биохимическому контексту и загруженной биологической функциональности. Опытно известные на сегодня возможности мутагенеза и общепринятые оценки возраста Земли не позволяют считать гипотезу самопроизвольного образования высших таксонов дарвиновским отбором, наложенным на стохастические мутации, правдоподобной. Что касается опытов Шапошникова и подобных им экспериментов, они лишь показывают, что потенциал искусственного отбора намного превосходит возможности естественного (см. здесь).
Я, конечно же, признаю возможность видообразования, однако настаиваю на том, что спонтанное появление функциональной новизны (достаточно большого количества функциональной информации), выражающееся, в частности, в особенностях морфологии (unique body plans):
Я, конечно же, признаю возможность видообразования, однако настаиваю на том, что спонтанное появление функциональной новизны (достаточно большого количества функциональной информации), выражающееся, в частности, в особенностях морфологии (unique body plans):
- исключено, если исключить возможность управления эволюцией (что, например, справедливо для классического дарвинизма и неодарвинизма);
- и, во всяком случае, ограничено сверху по количеству функционально-специфической информации, способной спонтанно генерироваться с учетом ограниченных вероятностных ресурсов биосистем.
Реальная картина относительно эволюции живых организмов представляется нечеткой. Филогенез имеет место, но в силу указанных причин ограничен низшими таксонами (количество спонтанно генерируемой функциональной информации, ответственной за биологическую новизну, ограничено). Чрезвычайная сложность организации имеющихся биосистем, а также ограниченность вероятностных ресурсов Земли дают основание предполагать, что фазовое пространство биосистем является архипелагом, а не континентом и что, следовательно, эволюция отдельных таксонов ограничена островами функциональности, разделенными областями хаоса. Таким образом, мне кажется, правильнее говорить не о едином филогенетическом графе, а о множестве графов, часть из которых, к тому же, вследствие эпигенетических эффектов, вообще говоря, не являются деревьями (см. также здесь).
Клеточные автоматы
В середине 20 в. фон Нейманом была предложена модель биологических систем в виде так называемых клеточных автоматов — регулярных двумерных структур, состоящих из ячеек, с наборами возможных состояний и правил по переводу ячеек из одного состояния в другое. Оказывается, существуют такие условия, при которых ячейки ведут себя согласованным образом, формируя устойчивые конфигурации. Эти конфигурации ячеек, находящихся в определенном состоянии, так называемые глайдеры (от англ. glide - "парить"), могут двигаться по пространству как единое целое. Известно также, что клеточные автоматы обладают свойством Тьюринг-эквивалентности, то есть могут выступать в роли универсальной модели вычислительного процесса. Поразительным при этом является достаточная простота и небольшое число исходных правил. Но и здесь мы все же имеем первоначальную информационную загрузку в виде установления правил (протокола обмена информации между ячейками) в дополнение к физико-химическим ограничениям реальных систем, моделирующихся клеточными автоматами. И снова подчеркнем принципиальную разницу между правилами, налагаемыми извне, и ограничениями физического мира. Ограничения, которые мы привыкли именовать законами природы, не могут выступать в роли правил.
Статистический аспект абиогенезных гипотез
Если я не ошибаюсь, еще Лейбниц говорил, что математическая грамотность проявляется в умении производить приближенные вычисления. Об этом же часто говорил мне мой дед со ссылкой на работы академика А.Н.Крылова [Крылов 1950] (о методах приближенных вычислений см. также [Меркулова & Михайлов 2007]). Когда я еще учился в школе, была произведена компьютеризация кафедр Брянского института транспортного машиностроения, в котором Игорь Алексеевич проработал доцентом много лет и где подготовил десятки инженеров. Я помню, как он сетовал на то, что расчеты в студенческих работах производились с чрезмерной точностью, не имеющей физического смысла вследствие приблизительности исходных данных. Скажем, значения модуля упругости дается с точностью до двух значащих цифр, а ответ приводится с точностью до шестнадцати, ибо "так посчитал компьютер".
Почему я сейчас об этом говорю? Дело в том, что аналогичную ошибку совершают многие специалисты - сторонники абиогенеза. Согласно абиотической гипотезе происхождения жизни, гипотетические протобиологические структуры возникли спонтанно в некоем первобытном океане. Однако при этом из виду упускается очевидная несостоятельность данной гипотезы с точки зрения статистики.
Рассмотрим некоторую функционирующую систему, например, двигатель внутреннего сгорания. Функционирование двигателя зависит от ряда параметров: от температуры масла, давления в поршнях, последовательности зажигания, процентного состав горючей смеси и пр. С увеличением числа параметров произвольной системы число теоретически возможных комбинаций их значений экспоненциально возрастает, тогда как размер областей фазового пространства, соответствующих рабочим режимам, быстро уменьшается. И в определенный момент оказывается, что вероятность того, что параметры системы окажутся в подобном функциональном островке, практически равна нулю.
Можно показать, что вероятность возникновения динамически устойчивой автономной структуры, способной самовоспроизводиться, поддерживать гомеостаз, выполнять простейшие метаболические реакции и реагировать на стимулы, на много порядков ниже оптимистической пороговой вероятности реализации любого правдоподобного сценария спонтанных и закономерных физико-химических взаимодействий в пределах Земли без участия интеллекта.
Указанная оптимистическая пороговая вероятность составляет 1 из 1070, или 10-70 и выведена из расчета принятых сегодня оценок возраста планеты (4.5 миллиарда лет, или 1017 секунд), максимально возможного числа атомов, могущих вступить в реакцию в земных условиях (1040) и верхней границы скорости возможных химических реакций (1013 реакций в секунду) [Abel 2009b].
Таким образом, гипотезы происхождения жизни, характеризующиеся независимо вычисленными вероятностями, меньшими 10-70, являются неправдоподобными, а сами вероятности должны считаться практически нулевыми. Рассуждения о вероятностях физико-химических взаимодействий, меньших порогового значения, лишены физического смысла и не имеют под собой никаких практических оснований.
Интересно отметить, что, согласно некоторым новейшим работам по биохимии, в опытах с бактериями установлено, что вероятность нахождения слепым поиском функционального белкового домена оценивается 10-77. Иными словами, в среднем функциональной является всего 1 аминокислотная последовательность из 1077. Таким образом, вопрос о переводе доменов в изначально функциональное состояние нетривиален, поскольку возможности спонтанных и закономерных процессов, протекающих в земных условиях, статистически исчерпываются.
Можно показать, что вероятность возникновения динамически устойчивой автономной структуры, способной самовоспроизводиться, поддерживать гомеостаз, выполнять простейшие метаболические реакции и реагировать на стимулы, на много порядков ниже оптимистической пороговой вероятности реализации любого правдоподобного сценария спонтанных и закономерных физико-химических взаимодействий в пределах Земли без участия интеллекта.
Указанная оптимистическая пороговая вероятность составляет 1 из 1070, или 10-70 и выведена из расчета принятых сегодня оценок возраста планеты (4.5 миллиарда лет, или 1017 секунд), максимально возможного числа атомов, могущих вступить в реакцию в земных условиях (1040) и верхней границы скорости возможных химических реакций (1013 реакций в секунду) [Abel 2009b].
Таким образом, гипотезы происхождения жизни, характеризующиеся независимо вычисленными вероятностями, меньшими 10-70, являются неправдоподобными, а сами вероятности должны считаться практически нулевыми. Рассуждения о вероятностях физико-химических взаимодействий, меньших порогового значения, лишены физического смысла и не имеют под собой никаких практических оснований.
Интересно отметить, что, согласно некоторым новейшим работам по биохимии, в опытах с бактериями установлено, что вероятность нахождения слепым поиском функционального белкового домена оценивается 10-77. Иными словами, в среднем функциональной является всего 1 аминокислотная последовательность из 1077. Таким образом, вопрос о переводе доменов в изначально функциональное состояние нетривиален, поскольку возможности спонтанных и закономерных процессов, протекающих в земных условиях, статистически исчерпываются.
Тот же принцип установления порогового значения вероятности в применении к нашей вселенной в целом при рассмотрении максимального числа планковских состояний с момента Большого Взрыва дает величину порядка 10-140..10-150.
Распознавание интеллектуального воздействия
Можем ли мы исходя лишь из анализа имеющейся конфигурации с достаточной степенью достоверности ответить на вопрос, имел ли интеллект отношение к ее возникновению? По всей видимости, да, можем. Для этого нам необходимо статистически исключить факторы случайности и закономерности. Согласно Уильяму Дембскому, это справедливо в случаях, когда имеется достаточно сложная независимо заданная спецификация и вероятность реализации данной конфигурации чрезвычайно мала (ниже порогового значения, о котором мы упоминали ранее). Некоторые примеры систем, для которых распознавание интеллектуального воздействия имеет положительный результат, таковы:
- Барельеф в местечке Маунт Рашмор в США (рис.8);
- Мегалитический архитектурный комплекс Стоунхендж в Англии (рис.9);
- Цифровые замки, распознавание кибератак, captcha-тесты и пр.
 |
| Рис.8. Барельеф в Маунт Рашмор, Южная Дакота, США. |
 |
| Рис.9. Стоунхендж, Великобритания. |
Задание независимой спецификации эквивалентно заданию рабочих областей в фазовом пространстве параметров. В случае биосистем островки параметров, как мы видели, чрезвычайно малы по сравнению с размерностью фазового пространства. По всей видимости, биофункциональность рассыпана по фазовому архипелагу, а не представляет собой один непрерывный континент, чего мы уже касались. Для перехода с одного островка к другому необходима комплексная перестройка имеющихся систем организма, тем более сложная, чем выше организация вида. Как мы уже видели, следует, таким образом, говорить не о едином филогенетическом древе, а о множестве графов, каждый из которых соответствует достаточно малым изменениям характеристик биосистем (в пределах заданной морфологии, в рамках высших таксономических единиц).
Спонтанная генерация принципиальной функциональной новизны статистически практически исключена в рамках физико-химических взаимодействий, реализующихся в земных условиях. Макроэволюция же по Дарвину предполагает самопроизвольную генерацию большого количества генетических предписаний, что исключено не только в силу статистического неправдоподобия, но и вследствие инертности материи по отношению к целеполаганию. Бесспорно, спонтанная коррекция ошибки и соответствующее увеличение информации не могут быть исключены.
- Скажем, Вы хотите отправить другу электронное сообщение "передай бабушке, что я приеду в 5 вечера", но случайно допускаете в нем ошибку, написав "приеду в 6 вечера". Получив послание, Ваш друг, также случайно исправляет ошибку, сообщая Вашей бабушке, что Вы приедете в 5 вечера.
Однако на практике в подобных случаях всегда имеется изначальное сообщение достаточной длины, составленное с использованием априорно установленных алфавита, синтаксиса и семантики.
То, о чем мы говорим, связано с так называемой теоремой о "бесконечных обезьянах". Эта теорема утверждает, что вероятность, с которой хотя бы одна обезьяна из стада случайно набьет текст "Гамлета" (порядка 130 тыс. символов) на компьютере, настолько мала, что в любых практических расчетах должна полагаться нулю.
То, о чем мы говорим, связано с так называемой теоремой о "бесконечных обезьянах". Эта теорема утверждает, что вероятность, с которой хотя бы одна обезьяна из стада случайно набьет текст "Гамлета" (порядка 130 тыс. символов) на компьютере, настолько мала, что в любых практических расчетах должна полагаться нулю.
- Точнее, если размер стада равен числу атомов в видимой части Вселенной (порядка 1080) и если при этом каждая обезьяна будет бить по клавишам со скоростью 1000 ударов в секунду, вероятность того, что хотя бы одна набьет текст шекспировской трагедии до тепловой смерти Вселенной, меньше 10-183000.
- Интересно, что объем представления 130000 символов в ряде современных языков программирования равен приблизительно 130 килобайт (из расчета 1 байт на символ). Для сравнения, объем генома человека составляет около 3 гигабайт. Предположение о том, что ДНК вообще и ДНК человека, в частности, содержит много "мусора", на данный момент уже практически опровергнуто. Участки кода, которые до недавнего времени было принято считать избыточными, на самом деле выполняют регуляторные функции в процессе синтеза белка. В этом смысле достаточно упомянуть тот факт, что хотя непосредственно белок-кодирующие участки составляют лишь несколько процентов всего объема генома, транскрипции для получения РНК подвергается почти весь геном.
- Заметим также, что, основываясь на приведенных аргументах, теория Дембского распознавания интеллекта устанавливает практический порог в 500 бит при наличии независимой спецификации системы. Утверждается, что если количество специфической информации превышает 500 бит, можно по статистике практически исключить закономерные и спонтанные факторы. Пороговое количество информации в 500 бит, ассоциированной с таким образом специфицированными зонами фазового пространства параметров системы, соответствует 72 ASCII символам осмысленного текста. Осмысленность любого текста большей длины явно свидетельствует о разумном его происхождении. В то же время известно, что наименьший объем простейшего генетического кода (вирусы) составляет порядка 7-9 килобит.
Я убежден в том, что настоящее творчество, приводящее к появлению достаточного количества функциональной новизны, не способно проявляться лишь под действием комбинаций спонтанных и закономерных факторов (подробнее о функциональной информации см. здесь). Фактически спонтанность и закономерность – стороны одного и того же: ограничений, задающихся действием законов природы, ведь закономерность есть та же случайность, только с вероятностью, стремящейся к 1.0. Творческий комбинаторный взрыв есть следствие третьей категории аристотелевой причинности: то есть выбора из возможных вариантов и связанных с этим выбором правил, наложенных поверх физико-химического «субстрата». Таким образом, жизнь не исчерпывается химией, хотя и включает ее, о чем говорили Макс Борн [Борн 1963] и Нильс Бор еще в середине XX века (cм. здесь и здесь):
Жизнь = Химия + Кибернетика.
Данное утверждение перекликается с античным тезисом о том, что жизнь представляет собой искусство. Сегодня мы можем по-новому это осмыслить, рассматривая творчество как процесс наделения систем функциональной информацией.
Эмпирическая наука говорит нам о том, что управление и сложная функциональность всегда есть следствие действий ЛПР. Но что же такое интеллект?
Мы можем ответить на этот вопрос феноменологически. Интеллект — это то, что сообщает функциональность сложным системам. Такое определение сродни определению гравитации как того, что обеспечивает притяжение тел, обладающих массой. Наше незнание природы гравитации не останавливает научную мысль. Мы можем измерять ее и учитывать ее эффект при построении научных моделей. Точно так же, незнание природы интеллекта не помешает привлекать его в наши модели. Так, по сути дела, и происходит в различных областях науки и техники, где осуществляется практическое распознавание интеллектуальной активности (см. некоторые примеры). Биология стоит особняком только лишь вследствие очевидных мировоззренческих ориентировок большой части ученых. Кибернетика является тем общим, что имеется в искусственных системах обработки информации и в живых организмах, из чего мы в полном соответствии с внутренней логикой научного метода делаем индуктивный вывод об артефактности биосистем.
Что такое интеллект?
Эмпирическая наука говорит нам о том, что управление и сложная функциональность всегда есть следствие действий ЛПР. Но что же такое интеллект?
Мы можем ответить на этот вопрос феноменологически. Интеллект — это то, что сообщает функциональность сложным системам. Такое определение сродни определению гравитации как того, что обеспечивает притяжение тел, обладающих массой. Наше незнание природы гравитации не останавливает научную мысль. Мы можем измерять ее и учитывать ее эффект при построении научных моделей. Точно так же, незнание природы интеллекта не помешает привлекать его в наши модели. Так, по сути дела, и происходит в различных областях науки и техники, где осуществляется практическое распознавание интеллектуальной активности (см. некоторые примеры). Биология стоит особняком только лишь вследствие очевидных мировоззренческих ориентировок большой части ученых. Кибернетика является тем общим, что имеется в искусственных системах обработки информации и в живых организмах, из чего мы в полном соответствии с внутренней логикой научного метода делаем индуктивный вывод об артефактности биосистем.
Наконец, тот факт, что математика как порождение нашего интеллекта и, так сказать, квинтэссенция формализма позволяет нам до некоторой степени формализовать реальность, тоже указывает на изначальный замысел, стоящий за самой физической реальностью [Abel 2011].
Литература
- Макс Борн (1963), Физика в жизни моего поколения. М., Иностранная лит-ра.
- К.Виолован (2004) "Анти-Перах: Слепая случайности или... Слепая случайность?"
- М.Ичас (1994) "О природе живого: Механизмы и смысл", Москва, Мир.
- Ю.Л.Климонтович (1996). "Введение в физику открытых систем", Москва, Изд-во "Физика".
- А.Н.Колмогоров (1965), "Три подхода к определению понятия 'количество информации'", Пробл. передачи информ., 1:1, 3-11.
- А.Н.Крылов (1950), "Лекция о приближенных вычислениях", Гос. изд-во тех. литературы, Москва, Ленинград, 1950, с.9-18.
- Н.Н.Меркулова, М. Д. Михайлов (2007), "Методы приближенных вычислений", Томский гос. ун-т, 2007.
- А.Финкельштейн, O.Птицын (2002) "Физика белка: Курс лекций", Москва, Книжный дом "Университет".
- David L. Abel (2011), The First Gene: The Birth of Programming, Messaging and Formal Control, LongView Press Academic: Biolog. Res. Div.: New York, NY.
- David L. Abel (2009a), The Capabilities of Chaos and Complexity, Int. J. Mol. Sci. 2009, 10, 247-291; doi:10.3390/ijms10010247.
- David L. Abel (2009b), The Universal Plausibility Metric (UPM) & Principle (UPP). Theoretical Biology and Medical Modelling, 6:27.
- D. Axe (2010a), The Case Against a Darwinian Origin of Protein Folds, Biocomplexity Journal.
- D. Axe (2010b), The Limits of Complex Adaptation: An Analysis Based on a Simple Model of Structured Bacterial Populations. Biocomplexity Journal.
- D. Axe (2011), Correcting four misconceptions about my 2004 article in Journal of Molecular Biology, Blog post, Biological Institute website.
- M. Behe (1994), Darwin's Blackbox: The Biochemical Challenge to Evolution.
- M. Behe, Self-Organization and Irreducibly Complex Systems: A Reply to Shanks and Joplin, Philosophy of Science 67 (March 2000), University of Chicago Press, August 31, 2000.
- M. Behe (2007), The Edge of Evolution: The Search for the Limits of Darwinism.
- M. Behe, W. Dembski, S. Meyer (2010), Science and Evidence for Design in the Universe. Proc. of the Wethersfield Inst., vol.9., Ignatius Press, 2010.
- E. Borel.Probabilities and Life, Dover, 1962.
- C. Darwin (1859), On the Origin of Species.
- W. Dembski (2007), No Free Lunch: Why Specified Complexity Cannot be Purchased without Intelligence , Rowman and Littlefield Publishers, 2007.
- W. Dembski, J. Wells (2007), The Design of Life.
- W. Dembski, R. Marks (2009). Conservation of Information in Search: Measuring the Cost of Success , Systems, Man and Cybernetics, Part A: Systems and Humans, IEEE Transations , Sept. 2009, 39(5), pp. 1051 - 1061.
- R. Deyes (2011), Proteins Fold As Darwin Crumbles. UncommonDescent.com, 2011.
- Gauger A.K., Ebnet S., Fahey P.F., Seelke R. (2010) Reductive evolution can prevent populations from taking simple adaptive paths to high fitness. Biocomplexity Journal, 2010(2):1-9.
- F. Glover and M. Laguna (1997). Tabu Search. Kluwer, Norwell, MA.
- B. Huberman, J. Mihm, C. Loch, and D. Wilkinson. Hierarchical Structure and Search in Complex Organizations, Management Science, Vol. 56, 831-848 (2010).
- S. Kauffman, Antichaos and Adaptation, Scientific American, August 1991.
- M. Kimura (1983), The neutral theory of molecular evolution. Cambridge.
- Z. Michalewicz, D.B. Fogel (2004), How to Solve It: Modern Heuristics, Springer.
- L.E. Orgel, 1973. The Origins of Life. New York: John Wiley, p.189.
- I. Prigogine, I. Stengers (1984), Order out of Chaos: Man's New Dialogue with Nature, Heinemann, London.
- J.F. Reidhaar-Olson, R.T. Sauer (1990), Functionally acceptable substitutions in two alpha-helical regions of lambda repressor , Protein Science, 1990;7(4):306-16.
- UncommonDescent.com.
- J. Wells, The Myth of Junk DNA.
- Wikipedia.org: сетевая энциклопедия.
No comments:
Post a Comment